I talked before about the challenges in supporting causes like Ferguson, where the best work is being done spontaneously and you have very little information. It turns out I do have a little bit of a connection- a good friend’s little brother goes to college in St. Louis, and he has a professor he considers a local expert on the subject of activism in St. Louis. I realize that readers as smart and informed as my own will give “blogger’s friend’s little brother’s professor” very little weight as a source. But sharing this information is better than not, so here it goes:
So after a bunch of back and forth with Bob Hansman, a professor here who is probably the person in St. Louis I trust the most to know how various charities affect people on the ground here, we decided that the best place to donate is the United Way of Greater St. Louis’s Ferguson Fund.
If you’re curious, I looked at various people on the ground and activist groups to see how to (or if we should) get them money. The problem was that many of these groups are not super active or transparent and that they advocate their own solutions to the complex issues at play here. For example, the most recent cause one of these groups organized for was a $15 minimum wage for fast food workers.
I wanted to send out a recommendation that was more broadly applicable. United Way seems to be pretty transparent and post a lot of updates on where this money is going. They also are supporting independent groups who are trying to figure out how to solve the problems in Ferguson, like the Ferguson Commission appointed by the governor. Hard not to get behind that.
I found this surprising. United Way is often held up as everything that’s wrong with charity: a big, lumbering organization more concerned with their own status than the people they are helping. And yet, they seem to be doing good work here, including supporting more nimble organizations. I think I will be putting some money towards this.
Normally when I’m investigating something I like to read well regarded books on both sides, in the hopes that the ignorance will cancel out. Finding a suitable counterpoint to Health At Every Size is hard, because its opposition is “everyone in the world”, and there has been no selective pressure to elevate the actual science away from the shame and aesthetic preferences.
For example, I spent a little bit of time on r/fatlogic which, as decisions go, was not my best ever. r/fatlogic frames itself as a criticism of horrible “fat logic”- things like “700 pounds is no less healthy than 200 pounds. Possibly healthier.” This is not a great start. I have a deep personal understanding of how frustrating it is when people are wrong, but I have found I am happier and a better person when I say “yup, wrong”, and then move on with my day. For the truly awful I might e-mail a friend making fun of it (thanks, Rachel!). Forming a whole club around criticizing people, especially people that are already having a pretty tough time in life, is bad for everyone.
r/fatlogic is even worse than that, because it has an extraordinary case of the cowpox of doubt. Wrong people keep insisting body fat is independent of calories consumed and exercise? Well then body fat must be solely dependent on calories consumed and exercise, and anyone who suggests it is affected by anything else is a fatty fat fathead making excuses for their fat. They are literally denying the possibility of individual variability in the translation of external environment into physical state. For bonus points, they invoke “but thermodynamics”, which is the same argument creationists use against evolution itself.
Here I tried several ways to explain exactly how wrong they were and how that was terrible, but then I decided to take my own advice and stop before I endorsed the hollow Earth theory. My current contender for an opposition book is Good Calories, Bad Calories, but I’m open to suggestions.
I read Health At Every Size and wrote a number of blog posts on it. Some are follow ups on science it mentions but didn’t not examine in depth, some are spot checks of its scientific claims. Neither set is comprehensive, but hopefully they are helpful.
HAES pre-check: What I thought about fat, food, and health before reading HAES.
Ghrelin: The Hunger and Lung Development Games: the discovery that the hunger hormone also plays a crucial role in fetal lung development inspires a pun I will find increasingly difficult to maintain as the series progresses.
Obesity, Blood Pressure, and Study Design: partially an examination of the link between obesity and blood pressure, but mostly an explanation of why the studies we’ve done so far are so unhelpful.
How did apples get to be the standard bearer for all that is good and healthy? In terms of nutrient per calorie, they’re not that good. 1 cup has 65 calories, 3 grams of fiber (12% RDA), and 5.7 mg vitamin C (10% RDA), and very small amounts of a wide range of other nutrients. Pears are just slightly better: 1 cup has 81 calories, 4g fiber, 5.7 mg vitamin C, and enough potassium to be noticeable. Meanwhile the same volume of grapes, so long derided as nature’s candy, have 104 calories, 1.4g fiber, 16.3 mg vitamin C, 22 mcg potassium. Almost everything apples or pears have trace amounts of, grapes have slightly more of.
Wasting their lives?
I wonder how much of this is because of the skin:pulp ratio. Produce skin tends to have a lot the bulk of the vitamins*. Plus grapes are more colorful, and color intensity is a shockingly good proxy for nutritional value in produce.
I also wonder how apples got such a sterling reputation without the benefit of a good marketing firm. My best guess is that they grow further north than most fruit and keep for much longer, and established cultural supremacy back when produce was scarce and fruit did not regularly fly.
Full disclosure: I was originally going to compare apples to iceberg lettuce, which I had previously seen described as nutritionally vacuous but easy to ship. But when I looked it up I discovered iceberg lettuce actually has a pretty good nutritional profile. 1 cup has 10 calories, 1 g fiber, 2.0 mg vitamin C, and 22.0 mcg potassium (22% RDA), and trace amounts of other stuff, which makes it strictly better than apples on a per calorie basis.
While we are at it: spinach does not have that much iron. It has a number of other vitamins and is very good for you, but the original reputation for iron-richness came from some guy putting the decimal point in the wrong place (source: some guy at a party 6 years ago). In fact the oxalates in spinach bind iron, making it harder to absorb.
Apples aren’t bad for you. If you want an apple, eat an apple. But if don’t want an apple and are trying to cajole yourself into it to make doctors keep their distance, consider grapes instead.
A chief contention of Health At Every Size (Linda Bacon) is that human beings can’t lose weight, so even if it would optimal for them to weigh less, there’s nothing to be done about it. Is this true? It’s hard to answer, because the question isn’t very well defined. Bacon admits there are things human beings can do to gain weight, and when they stop doing them, they sometimes lose weight. So if you’re doing those things, you probably can lose weight. And that people have a set range they can move around in healthily, based on diet and exercise, so if you’re at the top of your range now you could lose 20 pounds and still be okay.
On page 143, Bacon very strongly implies that twins maintain the same body weight even when they have very different activity levels, so weight is controlled by genetics. The studies she cites do show that when activity and diet is held constant, two unrelated people will have different health metrics. They also show that when two identical twins have different activity levels or diets, they will have different health metrics (including weight). Oh, and the combined sample size of both studies together is 35 sets of twins. This is where I started to get angry. I get using weak studies that strongly support your hypothesis. I get misleading people about what a study stays to support your hypothesis. But doing both is just…argh. I supported HAES. The actual prescriptions for food are basically the Michael Pollan diet (eat food, mostly plants), and motivation to retrain yourself to like real food rather than hyperprocessed crap. Those goals are good. Those goals are good even if they lead to weight gain, because under most circumstances produce is good for you and cheetos are not (although not all- this Captain Awkward post is full of people for whom carrots trigger intense digestive distress but hamburgers are safe and nurturing. I used to live off of pasta because anything else felt like eating death.)
I did check Bacon’s sources on the claim that people who lose dramatic amounts of weight tend to gain it back within 5 years. That appears to be true, at least in the studies she cited. And yet, she also cited studies showing that activity level affects weight. My explanation is that losing weight is not a thing you do. Your diet and activity level translate to a weight or body fat percentage*. If your current weight is different than that, it will move towards it. If you change your behavior, you will move towards the new translated weight. The translation appears to be a combination of genetics and perhaps past experience (she claims loss-and-regain cycles increase the set point. I’ve read that a lot of places, but at this point I neither trust her nor have the heart to investigate).
Impervious of weight to diet and exercise- depends a lot on what you mean. I will never look like Keira Knightley, but I will probably lose fat if I exercise more. Which I have just started to do after ceasing for a very long time because I was recovering from surgery, and will pause again when I have my next surgery, because the health harms outweighed the benefits. Fat is a proxy for health, but not the only measure of it.
Large amounts of fat are quite bad for you, but it’s unclear where that effect kicks in. The American aesthetic ideal is much lower that the healthy weight cut off, and may be actively unhealthy. In the normal range, diet and exercise have bigger health impacts than fat.
Our food supply is definitely fucked.
Fat people still don’t deserve to be shamed, especially under the guise of for their health. First because no one deserves to be shamed, but especially because shame is super bad for your health. It is intrinsically bad and it keeps people from seeking medical care, for both things related and unrelated to their fat. Stop doing it. People owe you neither their health nor their attractiveness.
*Weirdly, in my case it appears to be weight. I’ve had a shockingly consistent weight despite large changes in activity level and muscle mass.
The Washington Post has a cool infograph showing the results of a racial Implicit Association Test by state. I have a couple of problems with it.
First, the population sample is composed of people who went out of their way to take the test on the website, which is a long way of spelling “the population sample is invalid”. Second, the volume of use of implicit association in psychology seems to be driven more by a compelling story of what it means and psychologists’ sheer joy at having a thing they can measure, in a short lab test, with numbers and everything, than any actual evidence that the story is true.
But those are objections I would bring up to any presentation of this data. What is bothering me about this infograph in particular is a new and exciting variant on misleading axis choice. The Post chose blue to represent a lower IAT score (which, if implicit association’s claims are correct, means being less racist) and red to be a higher one. The lowest values are a darker blue, the higher a darker red, with a neutral value being white. At this point, blue and red are so tightly associated with Democrat/liberal and Republican/conservative that I think using them for anything else is manipulative. But having white as a middle color also strikes me as weird. Wouldn’t an even mix of the two be purple?
Worse, the “neutral” color does not rest on a score of 0, because that is the lowest score on the IAT. Instead white represents a score of 0.402, which is almost but not quite the middle of the range of state averages (0.341-0.456) It was chosen because it was the IAT score of the median state, Michigan. The overall effect is that a casual reading of the infograph would lead to conclude southeast and eastern states are racist and New England and northwest states are anti-racist. In fact, if you treat IAT score as a quantity, the most racist state is about 33% and 10 percentage points more racist than the least racist one. We don’t know what that corresponds to in actual behavior- does 10pp translate to a 1% difference in likelihood of hiring a black person, or 50%?- but that makes the color choices more misleading, not less.
For a long time now I’ve been trying to describe a hesitation I’ve had around EA. Outcome metrics are great. Outcome metrics are a huge improvement over “but look how much money we spent.” and “have you seen how sad this child is?“. And yet. My original stated concern was that over-reliance on metrics would drive us to focus on easy-to-measure outcomes over equally more* important hard-to-measure outcomes, or on known outcomes over more important unknown outcomes.*
Now I have a better analogy. Metrics are like nutritional labeling. Nutritional labeling is great when you want to decide between cheetos and soylent, or between soylent, mealsquares, and any one of their homebrew competitors.** But suppose I set a fiber quota for myself. The ideal way to do that would be to eat a variety of fruit, vegetables, beans, and nuts throughout the day, but that is super hard to keep track of. I either have to eat in exact serving sizes (forcing the continuous variable of hunger to the granular treatment of serving size) or calculate exactly how much I ate after the fact (a pain in the ass and/or impossible), and then look up how much fiber is in the food (ignoring any natural variation), write it down, total it up… and if it’s midnight and I’m short, eat a ton more food I may not want. Or I can pour a bunch of psyllium husks in a glass in the morning, check “eat fiber” off my todo list, and eat HoHos for the rest of the day.
Obviously the first choice is better overall, even if I ultimately end up with less fiber. But it is much harder to measure, in part because the benefits accrue over a wide variety of nutrients, whereas the psyllium and HoHos diet produces one big shiny number to trumpet in brochures. I think this is a problem in charity too. The Ugandan girls-club study I looked at last week had some outcomes that were both easy to measure and to value (spending), easy to measure but hard to estimate the value of (delayed marriage and childbirth), and kind of fuzzy to measure and of unclear value (age at which they do marry, as measured by proxy “when would you like to get married”). Luckily for that project the increase in girls’ income per unit NGO spending was almost as high as it was for pure vocational training, plus it had these social benefits, but suppose it had been 75% as good? Half as good? 10% as good? What is the cut off for being better than pure vocational training.
I’m solving this problem in my nutritional life by drinking a full serving of vitaminized protein powder*** mixed with chia seeds every day, plus whatever the hell I feel like eating. The almost-food frees up my stomach and brain to figure out what I especially need and seek that out, without fear I’m letting some other deficiency fester. This is startlingly similar to Holden Karnofsky’s (co-founder of GiveWell) suggestion that westerners focus on the problems of the 3rd world they are in a good position to fix (e.g. malaria), and let the locals do the rest. So I guess Effective Altruism has addressed this problem, it’s just that it addressed it by limiting itself, which is not the most emotionally satisfying answer but is something the world could do with more of.
BONUS FACT: EA and soylent have both found their home primarily with the rationalist community, and my rationalist friends (all of whom I met through EA) are simultaneously the most likely of anyone I know to drink soylent and to host communal dinners with secular grace.
*E.g. Food aid to the third world looks great measured by “people who stop starving in the short term.” We know now that this destroyed the local farming economy and left entire regions either starving or in ongoing dependence on 1st world aid.
**Of these, mealsquares have been the clear winner among my friends.
***Not quite the same as soylent because it lacks the fat, carbs, and fiber to be a meal replacement. This presents two slightly different problems. The lack of fat and sugar I feel fully prepared to make up for in the rest of my diet. But nutrients are digested differently depending on what other nutrients they are in proximity to. The chia seeds are attempt to get the benefits of protein x fiber.
I’ve talked a lot about controlling for confounding variables (also known as backing them out of your analysis), by which I mean, I’ve insulted several researchers for not doing it. And sometimes it is as obvious as I make it sound (controlling for smoking when studying coronary events). But sometimes it’s not. Eztra Klein has a great post about what it means to control for something. If black and white people receive equal sentences for using crack and powder cocaine the system is, in one sense, not-racist. But the fact that crack cocaine gets penalties so much higher than powder cocaine, and crack is more associated with black people, means that black people end up with much more jail time. If the sentencing discrepancy is based on good reasons (e.g. yes, murder should get more jail time than jay walking), it’s not racist. If the sentencing discrepancy exists entirely because crack is associated with black people, then it’s extremely racist, and the people who receive higher sentences for crack use are victims of racism against black people regardless of their race. Klein takes on racism in the context of crime. It’s true that you can almost close the criminal justice gap by controlling for things like income and type of offense, but those things are not necessarily independent of race. It’s an excellent piece and you should read it both for it’s excellent explanation of how statistical modeling works, and its commentary on race in America.
This comes up with obesity too. Someone went through and calculated life expectancy for each BMI* category (*moment of side eye for categorical analysis*), and found that while obesity (BMI >= 30) and being underweight (BMI < 18.5) were associated with excess deaths relative to normal weight, being overweight ( 25 <= BMI < 30), was not. First, I find it extremely telling that their description was “not associated with excess deaths”, when the more accurate description would be “associated with noticeably fewer deaths.” BMI-overweight people actually lived longer than BMI-normal people. Allow me to summarize the conversation that followed.
Fat advocacy groups: see, fat isn’t unhealthy. In fact, it’s the healthiest.
Pro-weight-loss groups: Bullshit. The study doesn’t account for things that make you thin and then kill you, like cancer, or anorexia.
FAs: Okay, so when a confound causes death and skinniness, it’s legitimate, but if it causes death and fatness** it’s an excuse to let ourselves go?
Team skinny: What’s the alternative, let you be fat? Let you feel good about being fat? I can’t talk about this with you until you’re ready to take it seriously.
What’s the lesson here? First, it’s to define your question very well. Do you care if literally changing someone’s skin color improves their life, or if there is an overall system in which one color consistently comes up as the loser? Is weight a good predictor of health outcomes, or is it a determinant of them? Designing experiments to tease out these variables is incredibly hard. Almost everything we know that affects weight also affects health. The exception is liposuction, and my understanding is it is not associated with any health gains, but then, it’s barely associated with weight loss. The situation with race, crime, and justice is even worse.
The second is that BMI is not only bullshit because it tosses a bunch of data away, but because the category boundaries are, at best, based on absolutely no data at all.
*Normally we would side eye use of BMI, but massive population studies are the one time it’s kind of okay.
**E.g. Type 2 Diabetes, sedentary lifestyle, poor diet\
This entry had better save someone’s life because WordPress ate the entirety of the first draft and half the second draft and it was not easy to recreate.
Health at Every Size claim: high blood pressure (hypertension) is not caused by fat, but by dieting (which, because fat people diet more, creates the illusion that fat causes high blood pressure. This is that confound thing we talked about last week).
My reading: The first source it cites does not quite say this: they say that current diet affects blood pressure, regardless of weight (which is actually even better news).* This study (PDF) purely retrospective and found both weight cycling and waist:hip ratio (a measure of fat gain around the waist, and due to the design of the study, a measure of fat itself**) to be significant (also, sample size was very small). The next 2 studies were in obese spontaneously hypertensive rats. Using animal models bred to have the disease you are studying is very common because it reduces the number of subjects needed to generate a statistically significant result, but if the model is suffering from a different root illness that merely causes identical symptoms (e.g. rats bred for leptin deficiency lose fat when given leptin, but leptin-sufficient humans do not). The last source is a book with a name like a perfume store.
Verdict: claim not proven.
HAES claim: hypertension is only associated with bad outcomes in thin people. Fat people with hypertension actually live longer.
My reading: This appears to be true, but all the studies were retrospective, none studies backed out smoking as a confound, and half were in men only. None looked for a dose-dependent effect, but dumped people in two or three buckets and compared outcomes within them. The technical term for this is categorical analysis (the alternative is numeric analysis), and it is almost always a bad choice. Here’s why:
START STATISTICS FIELD TRIP
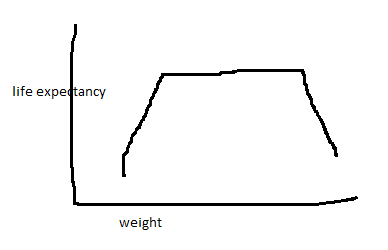
Let’s imagine that weight affects, and is the only thing to affect, longevity, and that it does so in an exceedingly simple way
Statistical analysis by MS Paint
Any idiot can draw the correct inference from this graph: being extremely overweight or underweight is bad, being anywhere in the middle range is equally good. Let’s imagine three naive scientists attempting to study the relationship between weight and longevity.
Scientist A gathers an equal number of people at every weight and sorts them into “low” or “high” weight, with the divider being the exact center of that trapezoid. They conclude weight has no impact on lifespan.
Scientist B draws the low/high line in the same place, but has slightly more extremely overweight people than extremely underweight people (which is what will likely happen in the real world, because 100 pounds overweight is a struggle, but 100 pounds underweight is dead). They conclude being overweight increases chance of death.
Scientist C gets and even distribution of weights, but draws the line to the left of the exact center. Their low-weight group now has a higher percentage of extremely underweight people than the high-weight group has of extremely overweight people. They conclude being underweight increases your chance of death.
And those were good intentioned scientists. A bad intentioned one can gather their data first and set cut off points after to prove almost whatever they want.
END STATISTICS FIELD TRIP
Verdict: After all the crap I’ve given HAES for misrepresenting studies, you might think I’m about to do that again. But actually I find it perfectly plausible that all the studies on weight and blood pressure use categorical analysis, because it’s incredibly common in medical studies. I really hope there’s a good justification for that, but the only one I’ve heard is that the math is easier, which might have made sense when everyone was doing statistics by candle light with a slide rule, but is hard to swallow now given the abundance of software packages that will do it for you. So I will tentatively accept HAES claim as true, but I don’t see how you can turn the claim into actionable suggestions, except to worry less, which is actually an extremely valuable suggestion in this context. So let’s go with “supported, but of limited usefulness.”
*This is your once-per-post reminder that someone who talks about weight or BMI and opposed to body fat percentage is already wrong.
**I spent several very confused minutes googling what “android” meant in context. I assumed the “android” referred to “women”, but android (when it doesn’t mean “obviously superior phone”) means “man-like”, which would make this an extremely unrepresentative study. Turns out that “android” modifies “obesity”, and it means fat growth around the waist and upper body (more often seen in men) as opposed to around the thighs, hips, and breasts (more often seen in women).
I have consciously decided to let the arguments for high protein diets pass me by, despite how convincing some of the research and underlying mechanistic claims sound. Humans have been cycling through high -protien, -fat, and -carb diets since we developed enough of a surplus to choose, and I’m pretty sure if any of them were that superior to the others it wouldn’t have been supplanted. I would say “everything in moderation”, but that is just a zen way of saying “Eat the correct amount. Idiot.” which I don’t think is very helpful.
HAES brings up a more constructive suggestion: eat fiber. Fiber* is usually left out of the macronutrient triumvirate, but so is water, and water is extremely useful. Off the top of my head: Fiber has the advantage of being natural**, because our ancestral foods had much more fiber than our current ones, yet its lack of calories is well suited to our current, couch-based, lifestyle. It evens out digestion of other nutrients, reducing the destructive sugar/insulin boom and bust cycle. It simultaneously treats diarrhea and constipation (if you drink enough water, which you should anyway). Have you ever wished you could eat a good thing and have it cancel out a bad thing?
Well, that might actually be true if the good thing is fiber, and the bad thing is not cocaine.
Here is where I meant to track down all the good things HAES says about fiber, but when I read it more carefully. I realized most of what it was praising was fruit, with the implication that fiber was the reason fruit is beneficial. The one actual fiber claim is that high-glyemic diets increase the risk of type 2 diabetes unless the diet is also high in fiber (five sources, all of which say nice things about fiber, one of which is actually about type 2 diabetes. The paper did conclude that fiber fights type 2 diabetes, but it was a post-hoc survey, which is a weak methodology). However, those other four sources also say pretty good things about fiber, just not the one the book claims they did. I think we are all on board with “fruit and whole grains” > “potato chips and soda”, although I’m severely disappointed that this of all books is backing that up with data on weight loss, rather than actual health outcomes.
But Dr. Wikipedia has many very specific nice things to say about fiber, and it cites much more relevant sources. Fiber increases micronutrient absorption (a little). Fiber increases nutrient absorption (more). Fiber fights inflammation. Basically, it does everything good and nothing bad. So while HAES didn’t lay out its case properly, fiber is definitely good, and I look forward to finding out who takes this too far and what the negative effects are.
*Definition of fiber varies a little from institution to institution, so for clarity: I am defining fiber as carbohydrates that are not broken down for energy in the human body. It’s worth noting that there are two contributors to whether an organism can digest food: its own enzymes, and the bacteria in its digestive track. Termites can’t naturally digest wood, but they house a bacteria that can.
**It’s easy to oversell “naturalness”, but when you have a mission critical system made of complex legacy code no one understands, sticking close to its original environment is a good default strategy.